Author(s): Ashraf Marzouk El Tantawi
PD-1 /PD-L1 is adoptive checkpoint mechanism, where PD-1 is an adopting its PD-L1 ligand activities by presence of specific helical kinase proteins in its compositions: _gamma common chains,_LNK” lymphocyte adaptor protein, _or SH2B adaptor _protein “tyrosine kinases, and _”SOCS” suppressor of cytokine signaling, that specified for controlling PD-1 and PD-L1 bindings activity through temporary resting the PD-L1 and consequently T-cells activities then transform incoming signals to exogenous processes in favor the of proper immune functions, But the permanent inhibition of PD-1 /PD-L1 binding is due to breaking down or inhibition in one or more of the adaptors helical proteins (lymphocyte adaptor protein, or SH2B) with the presence of SOCS” suppressor of cytokine signaling that will increase stability of binding without adopting and without activities, but cells survival can still exist through presence of tyrosine kinases , interferon stimulate kinases ,and gamma common.
Where, deficiency in lymphocyte adaptor protein, or SH2B adaptors with presence of “SOCS” suppressor of cytokine signaling in PD-1 will stimulate and increase binding stability of PD-1 to PD-L1 ligand (that. Can described as idle status of binding of PD-1 to PD-L1), that can lead to inhibition in T-cells activities. Cells death can started by broken antigens, ribosomal substances, OPA1 membrane and PD-L1 that will lead to releasing PD-1 due to its origin is related to Ser /Thr phosphorylation pathways. PD-1 biosynthesis is regulated by proper active Gamma-chain productions which produced and activated by JAK stat signals pathways which has the function of stimulating OPA1 synthetase oxidative functions in availability of NF-κB (that activate proper transcription, and binding via several intermediate steps, leads to an interaction with IκB kinase ) for activating IFN-gamma productions that will regulate and stabilize PD-1 , PD-L1, MHC-class-I, MHC class II, then SIRPα1 for TLR4 proliferation activities respectively.
ITSM is a conserved sequence of amino acids concluded in PD-1 cytoplasmic tail that found intracellularly in cytoplasm, but ITSM receptors can be found as gamma (on the surface of cells in PD-1), beta (in PD-1 within cells or on surface of cells) , or alpha (that its receptors found in PD-L1) and is carrying out the functions and responsibility of promoting and adopting their cellular activities including anti-inflammatory processes , that can transform transmitted signals to exogenous processes through the interactions of SHP-1 and SHP-2 that the absence “LNK” lymphocyte adaptor protein or SH2B adaptor protein From PD-1 will lead to missing the Adoptive functions in PD-1 that lead to continually binding of PD1 to PD-L1 that inhibit T-cells).
Absence or dysfunction of PD-1 activities (which has Antitumor and anti inflammation activities) can be due to the Incorrectly composition from helical proteins which build PD-1 that will be the result of promoting accumulations of more cytotoxic and increase inflammation, and survival of cancer invasion.
Gamma common and some other helical proteins such as: LNK lymphocyte adaptor protein, or SH2B adaptor protein 3, tyrosine kinases, and SOCS suppressor of cytokine signaling are regulating both globin and PD-1 biosynthesis, that both PD-1 and hemoglobin are having the function of recovering each other’s in proper conditions in vivo (in presence of proper NFκB & OPA1 enzymes), Where, in the proper biological condition the activations of NF-κB and OPA1 activities will expand cells survival.
Treatment of tumor Cancer chronic anemia, deficiency in immune efficiency, and deficiency in CD8 T-cells and NK cells activities. Some (chronic) Anemia can be results of chronic deficiency in: -γc gamma common, _LNK lymphocyte adaptor protein or SH2B adaptor protein 3, -SOCS suppressor of cytokine signaling, And deficiency in tyrosine kinases, With decreasing in NFκB with OPA1 activities lead to decreasing in IFN-gamma productions which regulate MHC-class-I, PD-1, PD-L1, MHC class II, and SIRPα1 activities.
Gamma-common (and other helical proteins such as: LNK lymphocyte adaptor protein or SH2B adaptor protein 3, tyrosine kinases and SOCS suppressor of cytokine signaling) are regulating both globin (globulin) and PD-1 biosynthesis, where both PD-1 and hemoglobin can recover each other.
It’s necessary in this study to understand the value of PD-1 Origin and their functions for cells prober survival and for proper cellular activities.
Several studies reported that The adapting immune cells activities is proceeded through antigens Adoptive activities for adopting anti-inflammations and endocytic proliferations, but adopting immune activities starts by Adoptive kinases and other kinase necessary for regulating activities which are activated by JAK STAT signalling pathways that are regulating most of active genes and subunits within cells and in exogenous that their activities will creat signals that reactivate neighbour cells created .
Firstly, the created signals started by the activities of ATPase, COX on pro-nutrients-mTOR molecules for producing long fatty acids chains followed by Ser/ Thr phosphorylation pathways for producing the main four kinds of kinases active molecules which will follow modifications by the effects of OPA1 anabolic enzymes for creating the three IFNs isoforms (the three Fatty acyl-CoA isoforms) where IFNs carry the necessary functions for adopting immune anti-inflammatory processes and proliferation specifically started by IFN-gamma synthesis ,which regulate antigen synthesis, T-cells activities, and NK cells functions through the activities and biosynthesis of PD1 and PD-L1.
The binding of programmed death-ligand-1 (PD-L1) to its Adoptive programmed death 1 ( PD-1) will inhibit temporary or reduce temporary, or adopt the exhausted T-cells activities for giving the priority to other immune cells to recover brocken cells, and antigens, that due to that binding will transform exogenous signals transmitted via Immuno-receptor Tyrosine-Based Switch Motif (ITSM) for reactivating exogenous IFNs (IFN-gamma), and glucocorticoids (GCs) functions for repairing Brocken tissue, cells, and their antigens, where the cytoplasmic tail of PD-1 contains two structural motifs, an ITSM and an immuno-receptor tyrosine the ITSM is a conserved sequence of amino acids concluded in PD-1 cytoplasmic tail (where, each active helical kinase has own receptor in its ligand “which formed from OPA1 oxidations” ) that found intracellularly in cytoplasm and is carrying out the functions and responsibility of promoting, adopting, and regulating anti-inflammatory activity , and transform transmitted signals to exogenous processes through the interactions of SHP-1 and SHP-2 for transforming and running activities with exogenous genes, and subjects of running and repair the brocken exhausted antigen and genes in immune cells [1].
Activation of IFNγ signaling leading to gene expression proceeds via activating cytokines through JAK-STAT pathway, followed by the tyrosine phosphorylation pathways. Class II MHC genes are inducible by IFNγ but generally not by IFNα/β that activating IFN gamma will proceeded via activating JAK STAT pathways followed by tyrosine phosphorylation (started by gammaoxidations and transcription). Activating gamma common and interferon stimulating receptor and adaptor kinases via JAK pathways will activate PD-1 synthesis which has cytoplasmic tail contains two structural motifs, an ITSM and an immunoreceptor tyrosine for running PD-1 activity [ 2].
The exogenous processes of cytoplasm tail can transform signals during the Adoptive binding of PD1 with PD-L1 during exhausted T-cells and NK cells for resting, reducing, and inhibiting their activities temporarily. Both PD-L1 and PD-1 are basically regulated by gamma common xhains and by interferon-gamma (IFN-gamma) that in the same time IFN-gamma can regulate Gamma-chain which regulate PD-1 synthesis (Gamma-chain produced from JAK and by IFN-gamma in mitochondrial matrix too). The reactivated OPA1 oxidative processes by cytokines is started by gamma-oxidation by effect of synthetase on gamma chains (which originally produced from Ser/ Thr phosphorylation signaling pathway then reactivated by jAK STAT pathway ) in availability of LPS that will produce fatty-acyl-CoA-synthetase, IFN-gamma, and glucocorticoid-gamma active subunits for regulating both PD-1 and PD-L1 biosynthesis productions.
During activating T-cells the PD-1 will be separated from its ligand PD-L1 and will be associated with created signals that can be transmitted into endocytic components for cellular biosynthesis depending on transmitted signals (including proliferation by TLR4 which associated with PD-L1) [3]. PD-1/PD-L checkpoint adoprive pathway is associated with substantial inflammatory effects, where the proper activity of PD-1/PD-L1 prevents inappropriate immune responses due to their proper physiological regulations, and due to the activiry of PD-1 which is promoting anti-inflammatory processes.
Due to substantial inflammatory effects will stimulate gamma common production that will stimulate mitochondria to activate the IFN-gamma production which will enhance MHC-class-I production which will be directed to nucleated cells for enhancing MHC class II and IFN-beta productions, that consequently will promote SIRPα1 and TLR4 synthesis (upon phospholipase effects) respectively And therefore will enhance PD-L1 synthesis and modifications.
IFN-gamma which promote T-cells activities is stimulated by gamma common chains productions from JAK state signaling Which enhance PD-1 productions that can be classified as type 1 cytokines which contain four helical protein necessary for PD-1.
Activities including checkpoint adopting kinase (which is so imp for adopting PD-1 function)
IFN-γ regulate T-cells activities, where can act on lipoproteins for producing gamma chains (which can be modified by JAK STAT) that with orher imp helixal kinases proteins will produce PD-1 (which will activate PD-L1 production ypon OPA1 enzymes activities) , or by activating the effector T cells soon after (but not before) of immune response , that PD-L1 expression is the major inducer at the transcription level [4,5]. It indicate that immune reactivation will be the result of reactivating gamma common kinase and other kinase that will stimulate IFN-gamma expression (upon OPA1 enzymes oxidations) which will reactivate swine leucocyte antigens (SLA) class I, class II expression, and will be the result of reactivating PD-1 again (through IFN-gamma cycles) that will be separated from its binding with ligand PD-L1 for running its own activity including anti-inflammation.
PD-L1/PD1 signals checkpoint adopting biosynthesis are associated with transmitting signals , that can accelerate the anti-inflammatory for analyzing tumor cells and inflammations, where, the proper active PD-L1/PD-1 signaling processes are characterized as one of the major mechanisms of tumor immune escape [6].
Signature of PD-1pos CD8+ T cells infiltrating MC38-derived murine tumor mass is having down regulated Drp1 activity and more fused mitochondria, compared to PD-1neg counterparts that signature of PD-1pos CD8+ T cells has strong fusion and connections to OPA1 enzymes activities for stimulating and running gamma-oxidation for producing active IFN-gamma (gamma-subunits) associated with active transmitting signals for activating producion of active acyl-CoA-beta, where both IFN gamma and beta having anti-inflammatory effect for analyzing tumor and adopting immune response (depending on adopting helical kinases productions and activation by JAK STAT pathway) cells activities [7].
That the reactivation PD-1 PD-L1 checkpoint Adoptive mechanism is typically promote the synthesis of the IFN-gamma “fattyacyl-CoA-synthetase” (gamma-oxidation) production followed by IFN-beta productions upon synthase effects.
Where during the active PD-1/PD-L1 checkpoint Adoptive mechanism will mask or hide glycolysis (but not inhibit) through fast hydrophobic amino acids synthesis (due to gamma oxidations) for stimulating IFN-gamma productions (where synthetase functions is pyrimidines synthesis for hydrophobic acids synthesis). It has been reported that PD-L1/ PD1 can alters T-cell metabolic reprogramming through inhibiting glycolysis and promoting lipolysis and fatty acid oxidation [8].
Where, to activate macrophages The glycolytic intermediate pyruvate can also metabolically control PD-L1 expression in macrophages through the BMP4/p-SMAD1/5/IRF-1 signaling pathway [9]. So, glycolysis will not inhibite during activation T-cells but at that time the periority will be for running gammaoxidation which will be the result of reactivating IFN-gamma productions, but purines synthesis is necessary for cells activities that is connected to pyrimidine synthesis for hydrophobic acids producions which is necessary for reactivating JAK STAT for producing active cytokines necessary for T-cells activities then for anti-inflammatory effects.
Also, activating PD-1/PD-L1 adoptive checkpoint will restores the imbalance of Th1/Th2 and Treg/Th17 cells [10].
That the PD-1/PD-L1 adoptive pathways will stimulate OPA1 enzymes activities for Re-activating gamma-oxidation which necessary for IFN-gamma productions then will be followed by beta-oxidation for pomoting swine leucocyte antigens (SLA) class I, and class II which will stimulated alpha-oxidations for SIRPα1 and TLR4 synthesis for proliferation and activate PDL1 synthesis for restore Th1/Th2 and treg/Th17[10] . Interleukin (IL)-1β, has emerged as a new tool in the therapeutic arsenal [11] that produced by beta-oxidations and by IFN-beta which have necessary functions in anti-inflammatory processes pathways.
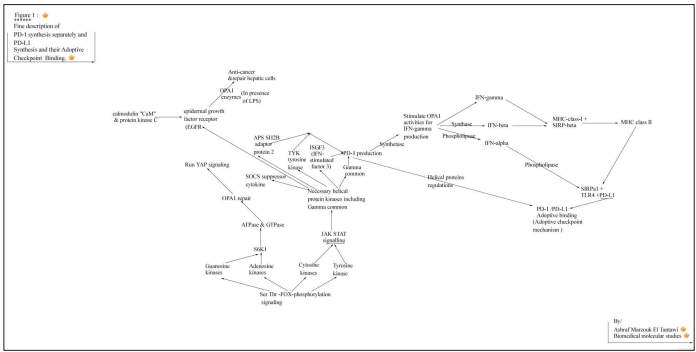
Figure 1

Figure 2
• Programmed cell death protein 1 (PD-1) programmed deathligand-1 (PD-L1)
• IFN-gamma
• Ser /Thr phospholipase signaling pathway,
• Tyrosine-Based Switch Motif (ITSM)
• SHP1 & SHP2,
• T-cells and NK cells,
• NFκB
• OPA1 Mitochondrial inner membrane
• The Janus kinase (JAK)-signal transducer and activator of transcription signaling
• BMP4/p-SMAD1/5/IRF-1 signaling pathway
• T-cells and naturel killer (NK) cells
• Th1/Th2 and Treg/Th17
• cytokines produced by Jack state signaling
• T lymphocytes _antigen-presenting cells (APCs)
• Tyrosine kinase 2 (TYK2.
• immunoglobulin
• Glycoproteins
PD-1/PD-L1 checkpoint has two main pathways:1) The 1st is binding which occurs for adapt immune activities by reducing or stopping exhausted T-cells activities whether for repair broken subunits and antigens, or for resting T-cells to be standby for any upcoming inflammatory signals, the IFN-gamma activity will be responsible for creating active signals (upon inflammation signals) that will activat separations of PD-1 fron its ligand “PDL1”, then PD-1 will be migrated to the cells surface to running anti-inflammatory activities.
The PD-1/PD-L1 Adoptive binding (not active signaling pathways) reduce or temporary inhibits the activities of exhausted T cells for repair their antigen and their endocytic Brocken genes and subunits. ‘‘Where, PD-1 is so related and connected to its ligand PD-L1, that in the absence of PD-1 will lead to absence of their main anti-microbial and anti-inflammation functions, but will lead to the appearance of the functions of PD-L1 separately without anti-inflammation function that will lead to tumor-infiltrating lymphocytes (TIL) abd tumor invasion, through increasing the function of proliferations with absence of PD-1. That it has been reported that bioinformatics study showed that PD-L1 expression was significantly associated with worse PFS of ovarian cancer’’.
The main function of PD-L1 is promoting and activating proliferation processes that is regulated by SIRPα1 and TLR4 synthesis upon phospholipase effects, where in case of absence or dysfunction of PD-1 (which has Anti-tumor and anti-inflammation activities) will promotes accumulation of more cytotoxic and terminally differentiated exhausted CD8+ T cells and will promote tumor-cells survival and cancer invasion [12,13].
The Inhibition of T cell antigen receptor (TCR)-driven stop signals (depended on continued interactions between PD-1 and its ligand, PD-L1) resulted in lower T cell motility, while the main function of PD-L1 is the regulation of increasing T-cells activities through proliferations while PD-1 regulate anti-inflammation through gamma common kinases and other helical cytokines including: tyrosine kinases , ISGF3 IFN-stimulated gene factor 3, and APS SH2B adaptor protein which will stimulate OPA1 for starting gamma oxidations and for producing IFN-gamma.
The programmed cell death-1 receptor (PD-1) is an immune adoptive checkpoint that can be defined as temporary inhibitor when bind with and linked to its ligands PD-L1, but can be permanent inhibitor in case of absence the APS SH2B adaptor protein from the composition of PD-1, Where that inhibition adoptive mechanism occurs by the adoptions by and APS SH2B adaptor protein to stimulate the binding by the presence of _SOCS suppressor of cytokine signaling as helical protein in PD-1.
The temporary binding between PD-1 and PD-L1 is only For repair exhausted T cells and for adopting immune processes , where, during reactivate PD-1 will be separated fron PD-L1 then migrated to the surface of nucleated immune effector cells. PD-1 is consider as the activator and regulator for PD-L1 which can be synthesis and regulated by the endocytic SIRPα1 and TLR4 in all nucleated cells.
Whereas, blockade of PD-1 enhances proliferation of memory B cells and expansion of virus-specific CD8 T cells during chronic simian immunodeficiency virus (SIV) infection in macaques. That indicate PD-1 enhance IFN-gamma production which enhance both MHC-class-I and MHC class II that will promote SIRPα1 and TLR4 for proliferation and promote PD-L1 production for proliferation too [14].
The biosynthesis of the programmed cell death protein 1 (PD-1) together with its ligand, programmed death-ligand 1 (PD-L1) stimulate OPA1 synthetase during stimulation for IFN-gamma synthesis (which regulated by synthetase enzyme) and then IFN beta will be formed (which depend on IFN-gamma synthesis) upon synthase effects , that promote the development (in case of proper conditions in vivo) , immunity evasion, but prevent the evasion of tumor cells and significantly prevent the survival of cancer, where PD-L1 only in the absence of PD-1 can prolong the survival of cancer disease.
That, PD-L1 on tumor cells is sufficient for immune evasion. Only PD-L1 (in absence of and deficiency in its PD-1) can increase evasion of normal and abnormal cells and tumor cells too, but in the presence and availability of PD1 will prevent tumor invasion, and will increase immune invasions, while PD-1 has the antiinflammatory effect and can be synthesis by gamma common chains [15].
The Janus kinase (JAK)-signal activator of transcription (STAT) plays critical roles in modulating of immune activities and cytokine receptors JAK/STAT signaling is essential for numerous developmental and homeostatic processes, including hematopoiesis, immune cell development, and gamma chain development. STAT5 as a direct modulator of PD-1. Where, the common gamma-chain (gamma c) cytokines IL-2, IL-7, IL-15, and IL-21, up-regulate PD-1. The common gamma chains cytokines play imp roles in PD-1 production and in all physiological processes , where PD-1 composition includes gamma common chains and four or five more helical proteins kinases necessary for regulating, adopting and activating proper activity of PD-1 functions [16,17].
Also, the Common γ-Chain Cytokines IL-2, IL-7, IL-15, and IL-21 Induce the Expression of Programmed Death. Reactivate gamma common will enhance synthetase oxidative function and will enhance PD-1, where both OPA1 synthetase and PD-1 will enhance the IFN-gamma productions which will regulate both MHC-class-I and MHC class II with contributions from synthade enzymes, then upon phospholipase effects will enhance SIRPα1, TLR4 and PD-L1 productions respectively [18] .
Recently, it has been found that endogenous glucocorticoids can induce selective and tissue-specific expression of PD-1 on NK cells. As gamma common regulate both PD-1 and enhance IFNgamma as also can enhance glucocorticoid gamma productivity where both IFN-gamma and GC gamma can recover and reactivate each other. And then GC can proof its functions for promoting and assisting IFN-gamma productions and functions for MHCclass-I productions and for recover PD-1, then for MHC class II synthesis (with synthase contribution) which will promote both SIRPα1 and TLR4 for PD-L1 synthesis respectively for necessary proliferation processes and consequently for reactivating T-cells and NK cells [19].
Both PD-1 and PD-L1 are necessary for adopting T-cells and immune activities and are depending on JAK STAT signaling (which mainly depend on Ser/Thr phosphorylation pathways), where There are four members of the mammalian JAK family, JAK1-3 and tyrosine kinase 2 (TYK2) which Are fully related to and depending on the productions of protein kinases PS/ TTK (thymine kinases) and PS/TCk, (cytosine kinases ) PS/ TAk (Adenosine kinases) and PS/TGk (guanosine kinases ) that produced from Ser /Thr phosphorylation signaling pathway that those four kinases are responsible for producing primary kinases which will be reactivated by JAK STAT pathway for producing their cytokines receptors including gamma common “and including other necessary helical protein kinases as tyrosine kinases and ISGF3 IFN-stimulated gene factor 3…. Etc” which included as helical protein in PD-1 compositions for adopting PD-1 activities and responsible for enhancing and re-stimulating IFN-gamma synthesis upon OPA1 synthetase effects that can run hydrophobic amino acids synthesis for running tyrosine phosphorylation pathways which plays imp roles in PD-1 activities [20].
The epidermal growth factor receptor (EGFR)is one of the receptor kinases that activated through JAK STAT signalling but originally produced by Ser /Thr FOX phosphorylation pathways as proteins kinases ( calmodulin “CaM” and protein kinase C “PKC” {Anna Llado, Paul[…]& Francesc Tebar (http://www.molbiolcell.org/ cgi/doi/10.1091/mbc.E07-05-0411) October 24, 2007} which produced from Ser/Thr phosphorylation exactly as Tyrosine kinases,, gamma kinases, S6K1… etc ) , where EGFR is having so imp roles in activating YAP signalling through stimulating and activating OPA1 enzymes activities for creating the three imp isoforms (gamma, beta, then alpha) necessary for running anti-inflammatory starting by producing acyl-CoAsynthetase(acyl-CoA-gamma) which will activate IFN-gamma (then IFN-beta, then IFN-alpha respectively) which is necessary for running anti-carcinoma and recover hepatic cellular activities. That “EGFR”-PI3K-PDK1 pathway regulates YAP signaling in hepatocellular carcinoma (EGFR)[21]. And the calmodulin (CaM) and protein kinase C (PKC) kinases regulate EGFR Cytokine receptors consist of multiple protein chains, where, are divided into two classes : Class I cytokines consist of four α-helices in a characteristic up-up-down-down configuration [22].
• γc gamma common
• SHP1 Src homology region 2 domain for adopt immune activities
• LNK” lymphocyte adaptor protein or SH2B adaptor protein 3
• ISGF3 IFN-stimulated gene factor “SOCS” suppressor of cytokine signaling
• Tyrosine kinases
Where, those gamma cytokines (including Gamma-chains) are able to promote PD-1 synthesis and stimulate OPA1 synthetase (in availability of NF-κB) activities for activating IFN-gamma (due to presence of ISGF3 IFN-stimulated gene factor) productions and activities, and then pyrimidine nucleotides synthesis for hydrophobic amino acids synthesis including Tyrosine (which necessary for P-Tyr phospho-tyrosine and JAK STAT pathway ) that can ré-stimulate, IFN-gamma, and for re-producing PD-1 (and regulate PD-L1) which looks like as class 1 cytokines that consists of four or five helical bundle proteins (concluding “LNK” lymphocyte adaptor protein or “SH2B” adaptor protein) for adopting activities by PD-1 [22].
PD-1 blockade does not have a significant effect on direct cell dependent lysis of leukemia, but PD-1 composirions has the role of stimulating and modulating IFN-gamma, Where PD-1 signaling modulates interferon-γ production [23]. The Stimulation of TLR4 in human CMFs upregulates PD-L1. Stimulation of TLR4 in human CMFs upregulates PD-L1 but poorly induce PD-1, That SIRPα1 and TLR4 are carrying the main function for proliferation respectively including PD-L1 activities regulated by phospholipase. where,SIRPα1 is promoting TLR4 biosynthesis [24, 25]. And, SIRPα1 regulated by MHC-class-I (which regulated by IFN-gamma ) and then by MHC Class II respectively [26].
TLR4 expression correlated with PD-L1 expression indicates a poor prognosis in patients with peripheral T-cell lymphomas. That both TLR4 and PD-L1 are essential for proliferations but not for anti-inflammation functions where absence of PD-1 will not give results of Antitumor or anti-inflammations [27].
And both TLR4 expression level was also positively correlated with the PD-L1 expression [28]. And also, the TLR4- and TLR7/8- medated are essential for upregulation of PD-L1 on plasma cells and DCs, respectively [29]. And, lipopolysaccharide via TLR4/ MyD88/NF-κB pathway promotes immune escape in pancreatic cancer [30].
Where presence of MyD88 stimulate IFN-gamma activity and consequently stimulate both PD-1 (for anti-inflammation) and PD-Ls, also will promote beta-oxidation by synthase, but NFκB will promote transcription Mitochondrial OPA1 oxidations that restimulate gamma-oxidation. Where, both NFκB and mitochondrial OPA1 contribute substantially to cell survival. NFκB and mitochondrial OPA1 can evaluate transcription processes and gamma, beta, and alpha oxidations needed for cells survival [31].
Also, the receptor programmed cell death 1 (PD-1) which considered as CD279 can alters T-cell metabolic reprogramming by inhibiting glycolysis and promoting lipolysis and fatty acid oxidation. that PD-1 (that contain helical APS SH2B adaptor protein 2 can adopt glycolysis for increasing lipolysis through stimulating OPA1 synthetase for modulating and regulating IFNgamma production by the presence of the helical protein [32].
“ISGF3 IFN-stimulated gene factor 3”) has roles of restimulating IFN-gamma activities throughout “ISGF3 IFN-stimulated gene factor 3” by stimulating synthetase activities which will show activating of lipolysis and fatty acids oxidations due to stimulation of OPA1 oxidative activities and will give the results of altering T-cell metabolic reprogramming and adopting anti-inflammatory activities due to the presence PD-1 activities.
V-ATPase family regulates lysosomal exocytosis and neutralizes with tumor-infiltrating lymphocytes for glioblastoma cells S6K1 modulate ATPase family which regulates lysosomal exocytosis and neutralizes with tumor-infiltrating lymphocytes for glioblastoma cells through their ability for repair ribosomal composition which are responsible for promoting GTPase for modulating OPA1 repair and recovery, that in case of reactivating the proper JAK STAT signaling specifically for producing necessary helical receptor kinases as: proper gamma common, SH2B adaptor protein 1, and ISGF3 IFN-stimulated gene factor 3 for reactivating PD-1 and for enhancing synthetase functions for IFN-gamma and for glucocorticoids-gamma (GC-gamma)which will modulate T-cells and immune anti-inflammatory activities [33].
IFN-γ has strong and diverse effects on various levels of hematopoiesis, it profoundly regulates blood cell production during immunologic challenges. And, Interferon-gamma modulates fetal hemoglobin synthesis in sickle cell anemia [34,35].
IFN-gamma modulate both of hemoglobin and PD-1, but gamma common chains regulate both PD-1 and hemoglobin production , then consequently there is a specific regulations between PD-1 and hemoglobin that are connecting and recovering each other and can enhance “stimulate” IFN-gamma productions
And in the same time Two gamma chains together with two alpha (which formed in mitochondria) chains constitute fetal hemoglobin, so gamma common regulate both PD-1 and globin synthesis (where both can recover each other), where PD-1 composition containing approximately 5 helical proteins:
1. γc gamma common,
2. SH2B adaptor protein,
3. ISGF3 IFN-stimulated gene factor 3,
4. TYK tyrosine kinase,
5. SOCS suppressor of cytokine signaling
As common gamma regulate IFN-gamma, GC gamma, and PD-1, as PD-1 can have the function and activities to recover and recover IFN-gamma, GC-gamma, and globin synthesis (in presence of NFκB & OPA1 enzymes) and consequently hemoglobin synthesis. During anti-inflammatory processes and cellular activities PD-1 will enhance (with adoption) OPA1-synthetase for promoting IFN-gamma productions that will produce first acyl-CoA synthetase (acyl-CoA-PD1-gamma) which upon synthase will produce PD-1-beta (or fatty-acyl-CoA-PD-1-synthase) which upon phospholipase effects will produce PD-L1 (fatty-acyl-CoAPD-1-alpha) which necessary for proliferation, and also also PD-1 can enhance JAK STAT signaling activity and OPA1 to promote and recover hemoglobin productions in the availabilities proper NF-κB and OPA1 proper activities.
Each proper kinase protein molecule has its own interacting receptor that modified and improved by JAK STAT signalling and by OPA1 enzymes activities, where OPA1 enzymes interact with kinases to produce their acyl-CoA isoforms that carry their own interacting receptors. Kinases proteins are the first tools produced firstly by Ser/ Thr phosphorylation pathways that will stimulate mitochondria enzymes to produce own acyl-CoA isoforms started by acylCoA-gamma production which will interact with own receptor for stimulating and promoting cellular processes with accurate adoptions (which regulated by their own helical kinases protein produced by JAK STAT signalling) including anti-inflammatory cycles and proliferation.
Heme is synthesized in a complex series of steps involving enzymes in the mitochondrion. The sythesis of heme involves multiple enzymatic steps process begins in mitochondrion by condensation of succinyl-CoA and glycine. The synthesis of hemoglobin begins with two distinct globin chains (each interacts with its own individual heme molecule) combine to form hemoglobin. One of the chains is designated as alpha, where the second chain is designated as “non-alpha” or gamma.
The fetus has a distinct alpha and non-alpha chain “gamma”. After birth, a different non-alpha globin chain, beta pairs with the alpha chain start to be formed. The Two gamma chains together with two alpha chains constitute fetal hemoglobin (HbF) that kinases protein received from breast feeding will be analyzed by synthetase for hydrophobic production and for gamma chains production which interact (stimulate) with OPA1 synthetase for acyl-CoA synthetase then will stimulate directly phospholipase effects for TLR4 synthesis for proliferation processes but after 6 weeks and more the beta kinase will be produced by creating glycogen (synthase synthase effects on transcripted gamma) which will be in a progressive scale for creating anti-inflammation tools, CD8 T-cells, and NK cells then the IFN-gamma will be synthesised that will promote SIRP-gamma, MHC-class-I and PD-1 (which regulated by gamma chains) then synthase will reactivate PD-1 to form acyl-CoA synthase that will run the flow of biological processes for TLR4 synthesis by phospholipase effects for increasing proliferations of plasma-membranes, for PD-L1 synthesis , for collagen synthesis, and blood platelets,where synthase enzyme can act on glycogen kinases for reactivating PD-1 function (to be separated from its PD-L1 binding mechanism ) and for reactivating PD-L1 for proliferation and for re-activating CD8 T-cells. As MHC-class-I, pd1 produced by IFN-gamma regulations as synthase enzyme will act on MHC-class-I and PD-1 to be modified (for PD-1-acyl-CoA-synthase protections) and to be migrated to nucleated cells for endocytic MHC class II production which will promote SIRPα1 and TLR4, and promoting PD-L1 synthesis upon phospholipase effects for proliferations processes [36].
Liver glycogen synthase (GYS2), a key enzyme in glycogen synthesis and play imp roles in activating PD-1 and then activating CD8 T-cell and NK cells. Also, glucose-6-P is involved in the regulation of heme-regulated eIF-2 alpha kinase, so, therefore glucose-6-P is playing imp roles in reactivating PD-1 and CD8 T-cell and NK cells (with still surviving the PD-1/PD-L1 Adoptive mechanism) [37, 38].
Activity and membrane association are regulated by Src-mediated tyrosine phosphorylation. NADH and FADH2 and then the TCA cycle are activated by the electron transport signal transduction which yield from JAK STAT signalling and from gamma-oxidation processes, that there is an absolute requirement for some G6PD activity, residues essential for coenzyme or substrate binding are rarely modified,as decreasing in JAK STAT pathway as will result in decreasing in G6PD-dependent as will results in decreasing in gamma common for PD-1 synthesis and will results in deficiency in composition of PD-1 helical protein kinases as will result of decreasing in globin synthesis and in heme too [39,40].
Gamma common chains are necessary for “non alpha” gamma necessary for fetus hemoglobin synthesis and development, and also in the same time are so necessary or PD-1 synthesis which are so imp for stimulating IFN-gamma production which is necessary for antigen re-synthesis too. Glucose-6-P allows maintenance and reduces oxygen radicals produced by cellular metabolism, and G6PD deficiency can cause neonatal jaundice and acute haemolytic anemia triggered by increased oxidative stress. where, glucose 6 phosphate promote alpha heme synthesis and glycogen synthase productions that allows anti-inflammation processes and promote PD1 re-activities (that stop the Adoptive binding of PD-1 with PD-L1 then will be separated ), and then will reactivate T-cells and NK cells [41].
Gamma common chains and helical protein kinases (from The Janus kinase “JAK” stat pathways) are necessary for “no alpha” gamma necessary for fetus hemoglobin synthesis and development , and at the same time are so necessary for PD-1 synthesis which are so imp for re-stimulating IFN-gamma production for antigen re-synthesis and reactivating T-cells and NK cells. So, gamma common (and helical proteins) regulate both globin and PD-1 biosynthesis, that PD-1 have the function of recover globin and vice versa (in presence of NFκB & OPA1 enzymes) and consequently hemoglobin synthesis.
Increasing in G6PD-dependent will be the results of increasing in gamma common for PD-1 synthesis and will results in increasing in composition of PD-1 helical protein kinases and will result of increasing in globin synthesis and in heme too. Glucose 6 phosphate promote alpha heme synthesis and glycogen synthase productions that will allows anti-inflammation processes and promote PD1 re-activities, then will reactivate T-cells and NK cells. Death of cells occur due to Brock in chromosome and in mitochondrial OPA1 that lead to inhibition in mitochondrial activities and in SIRPα1, TLR4 and in PD-L1 lead the cell death to release PD-1 due to PD-1 its biosynthesis is originally related to Ser/Thr phosphorylation pathways outside cells . Where, PD-1 enter the cell that will stimulate OPA1 activities for producing (gamma-subunits) IFN-gamma upon synthetase oxidations followed by beta and then alpha oxidations upon synthase and phospholipase effects respectively.
In brief, the helical proteins kinases that activated and produced by JAK STAT signalling are having the functions of regulating many active genes and imp subunits “including PD-1 function”, and their proper productions are able to protect cells from death, and also able to regulate most of cellular activities including proliferations.
The binding of PD-1 /PD-L1 is done due to two reasons : 1st/ is adopting and repair cells by translations & transcriptions will helping cells to re-exist again by recovering their necessary proliferation which done by SIRPα1, TLR4 and PD-L1, and will recover their protections by PD-1 which carry functions of anti-inflammations and re-stimulating IFN-gamma activities, 2nd/ both PD-1 and PD-L1 are recovering hemoglobin which catalyzed during anti-inflammations, where PD-L1 is formed in mitochondria (regulated by OPA1 synthetase and consequently by IFN-gamma) exact as hem when produced in Mitochondria , and globin is formed and regulated by gamma common which regulate and promote PD-1 synthesis too.
The value of the roles of functions of both PD-1 and PD-L1 are so imp for recovering their own cells and other cells too even after the death of their own cells that both PD-1 and PD-L1 will try to re-exist for healing other cells and hemoglobin too.
The Author declare that the research work has been studied and conducted in the absence of any commercial or financial relationships, that could be construed as a potential conflict of interest.